فصل هفت
الکتروفیزیولوژی قلب
سامانه پیشآهنگ و رسانش
- گره سینوسی دهلیزی
- گره دهلیزی بطنی
- تار های هیس
- انشعابات تار های هیس
- مسیر های آناتومیکی میانبر گره دهلیزی بطنی
قواعد پایه الکتروفیزیولوژی
- پتانسیل عمل غشایی
- دلالان های غشایی
- وابستگی ولتاژی
- وابستگی زمانی
- خودکاری
- خودکاری در مقابل تحریک پذیری
- سرکوب فرازش
- پتانسیل استراحت غشایی
- پتانسیل آستانه
- مرحله سرکشی(مقاوم)
- حالت سرکشی(مقاومت)
- سرعت رسانش
- عملکرد دروازه ای یاخته های پورکنژ
الکتروکاردیوگرام
- رابطه پتانسیل عمل با الکتروکاردیوگرام سطحی
- نظریه دو قطبی معادل
- ثبت الکتروکاردیوگرام
- کاردیوگرافی برداری
- رونوشت سازی از الکتروکاردیوگرام
- تجزیه و تحلیل الکتروکاردیوگرام
- اندازه گیری مجموعه P-QRS-T معمول
- تجزیه برداری
- انواع الکتروکاردیوگرام از پستانداران بالغ
الکتروکاردیوگرام غیر عادی
- نمودار های نردبانی
- آریتمی های قلبی(بدآهنگی های قلبی)
کاربرد بالینی الکتروکاردیوگرام
سامانه پیشآهنگ و رسانش الکتریکی
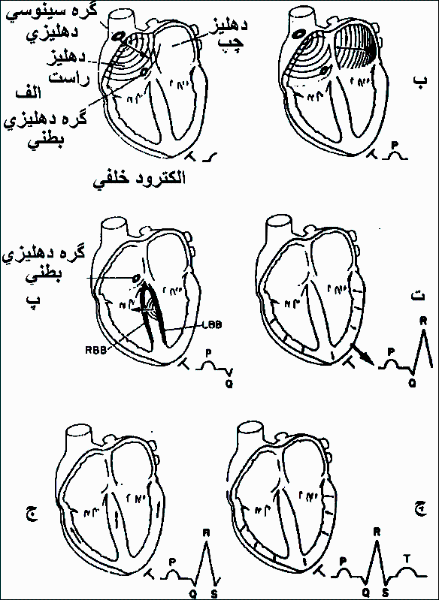
بافت های پیشآهنگ ویژه و هادی الکتریکی قلب عبارتند از گره سینوسی (سینوسی دهلیزی) ، گره دهلیزی بطنی، نوار هیس، انشعابات سمت چپ و راست از نوار هیس و شبکه محیطی پورکنژ(تصویر 1-7).
تصویر 1-7: سامانه رسانش قلب پستانداران.
بطور معمول یاخته های پیشآهنگ واقع در گره سینوسی دهلیزی مسئول آغاز تپش های قلبی هستند. اگر چه رشته های آناتومیکی خاصی که گره های سینوسی دهلیزی و دهلیزی بطنی را به یکدیگر مرتبط کرده و یاخته های دهلیزی که به لحاظ الکترو فیزیولوژیک برای رسانش اختصاص یافته اند بررسی و توصیف گردیده اند، رسانش در دهلیز اساساً از طریق یاخته های معمول عضلات دهلیزی قلب محقق می شود . رسانش فوق در طول محور طولی یاخته های فوق سریع تر از محور عرض آنها است. احتمالاً نحوه و ترتیب چینش رشته های عضلانی فوق علت هدایت سریع در طول مسیر های مقدم بر گره دهلیزی بطنی می باشد تا رشته های دسته مانند یاخته های اختصاص یافته . در هر دو جنس سگ و خرگوش ستیغ انتهایی و لبه قدامی سوراخ بیضی اصلی ترین خط سیر برای چنین رسانشی میان گره های سینوسی دهلیزی و دهلیزی بطنی بوده و اجازه فعالیت زودرس را به گره دهلیزی بطنی می دهد.
معمولاً تکانه های قلبی که به بطن می رسند ،بایستی با گره دهلیزی بطنی تقاطع یابند، ناحیه ای که بطور نسبی قابلیت رسانایی آهسته ای دارد. گره دهلیزی بطنی معمول ترین مسیر مابین میوکارد دهلیزی و نوار هیس می باشد . تکانه های برآمده از گره دهلیزی بطنی به رشته های تند رسانای پورکنژ نوار هیس می رسند و از طریق انشعابات نوار فوق به شبکه رشته ای پورکنژ بطن ها انتقال می یابند.
این سامانه مرکب، مسئول سمت گیری و انتشار تکانه های قلبی در طول قلب و در هر تپش آن می باشد . سامانه فوق موجب انقباضات پی در پی دهلیز و بطن شده و کل توده های عضلانی بطن های قلبی را بطور همزمان منقبض می نماید. قطر و سرعت هدایت الکتریکی رشته های عضلانی قلب پستانداران در جدول 1-7 آمده است .
جدول 1-7: قطر رشته و سرعت هدایت الکتریکی یاخته های عضلانی قلب پستانداران.
گره سینوسی
گره سینوسی در دیواره دهلیزی سمت راست و در محل اتصال سیاهرگ میانخالی قدامی با دهلیز راست جای گرفته است . این محل اتصال در سطح اپی کاردی بصورت شیار انتهایی و در سطح اندوکاردی بصورت ستیغ انتهایی تظاهر می نماید . گره سینوسی دهلیزی در سگ، انسان و چند گونه دیگر به شکل بیضی می باشد، ولی در اسب به شکل نعل اسبی با ساقهای مخروطی طویل و با شکل مشابهی در دیگر تک سمی ها دیده می شود . ساختار چندگانه آن در چندین گونه مختلف مشابه می باشد. یاخته های عضلانی طویل و ظریف ، با تارچک های عضلانی پراکنده که در یک شبکه غنی کلاژنی پیچیده شده اند این ساختار فشرده را تشکیل می دهند.
گره دهلیزی بطنی
این گره در بخش زیرین اندوکارد دهلیز سمت راست در قاعده دیواره بین دهلیزی دقیقاً در ناحیه قدامی دهانه سینوس تاجی واقع شده است . در سمت دهلیزی این گره شاخه هایی از عضلات دهلیزی را دریافت کرده و با نوار هیس در سمت بطنی امتداد می یابد. تار های این گره که از عضلات دهلیزی کوچکتر هستند الگوی منشعب و در هم تافته ای از رشته ها را در یک شبکه از بافت همبندی تشکیل می دهند. یاخته هایی متنوع موجود در این بافت قلبی اختصاص یافته، ویژگی های الکتروفیزیولوژیکی گوناگونی دارند .
نوار هیس
در فاصله مابین گره دهلیزی بطنی و نوار هیس ، یاخته هایی حضور دارند که در رشته های منظم تری جمع شده و تار های مشخص پورکنژ را می سازند. این رشته های پورکنژ تجمع یافته را نوار هیس می نامند . نوار هیس بطور میانی از گره دهلیزی بطنی واقع در اندوکارد دهلیز راست و در طول لبه پایینی دیواره بین بطنی عبور کرده و آنگاه به یک تک شاخه نواری سمت راست و شاخه های متعدد نواری سمت چپ منشعب می گردد.
انشعابات نوار هیس
نوار سمت راست هیس ابتداعاً منشعب شده و از نوار دیگر کوتاه تر است . شاخه های نوار سمت چپ در ادامه نوار هیس بصورت نوار پهنی است که بصورت زیراندوکاردی نزولی در سمت چپ دیواره بین بطنی کشیده شده و قبل از تشکیل شبکه پورکنژ محیطی به نوار های اصلی قدامی و خلفی تقسیم می گردد (تصویر 2-7).
ریخت شناسی رشته های موجود در انشعابات شبکه پورکنژ محیطی در نمای میکروسکوپ الکترونی بسته به ابعاد قلب در گونه های مختلف پستانداران متفاوت می باشد. در پستانداران کوچک (مثل گربه، خرگوش، خوکچه هندی، موش) این یاخته ها از انواع عضلات قلبی کارگر قابل تمییز بوده و تنها تفاوت یاخته های پورکنژ در نداشتن سامانه T می باشد . در پستانداران بزرگ (مثل سم داران ، والها) یاخته های پورکنژ بزرگ بوده و رنگ پذیری اندکی داشته و حاوی پروتئین های انقباضی ناچیزی می باشند. همچنین در پستانداران بزرگ و پرندگان شبکه پورگنژ بطور گسترده ای بداخل میوکارد بطنی نفوذ داشته ولی در پستانداران کوچک به لایه زیر اندوکاردی محدود می باشد . در مهرداران کوچک پست که قلبی کند تپنده دارند ، هیچ نوع سامانه هدایتی وجود ندارد. روند رسانش در یاخته های بزرگ با سرعت بیشتری انجام می شود (تصویر 3-7). این تطابق ریخت شناسی در قلب های بزرگ اجازه انتشار سریع تحریک را می دهد(مثل والها و سم داران) و از طرفی سرعت رسانش سریعی را برای ضربانات پرسرعت پرندگان فراهم می آورد . شمار گوناگونی از رشته هایی با ابعاد متوسط (رشته های انتقالی) به همراه تارهای پورکنژ انشعابات نوار هیس را می تواند دید.
تصویر 2-7: سامانه هدایتی(رسانش) بطن چپ در دو قلب مطعلق به سگ. سطح اندوکاردی با ید رنگ آمیزی شده . شاخه کلافه(دسته) سمت چپ(LB) چون ساختاری نوارمانند در زیر ریشه آئورتی در هر دو مورد تظاهر می نماید. LB به دو دسته قدامی(A) و خلفی(P) منشعب شده، بطوری که به بخش های عضلات پردی(پاپیلی) متصل می شوند. شبکه ای از تارهای پورکنژ بین دیواره ای در سطح بین دیواره ای انتشار یافته و توسط دو دسته تار اصلی همراهی می گردد. مابین رشته های پورکنژ دو قلب فوق پاره ای اختلافات موجود هست. آئورت(AO) و دیواره بین بطنی(S) نیز در شکل نشانگذاری شده است.
تصویر 3-7: سرعت هدایتی نسبی(نمودار بالایی) و بیشترین سرعت قطبش وارون (نمودار پایینی) در یاخته های گره سینوسی دهلیزی(SAN)، دهلیز (ATR) ، ناحیه گره دهلیزی(AN) ، گره(N)، گره-هیس(NH) ، نوار هیس(BH) ، انشعابات نوار هیس(B.B) تار های پورکنژ(PF) ، تار های انتقالی(TRAN) ، عضلات بطنی(VM) .
بتدریج از ابعاد این رشته های انتقالی کاسته شده و در ادامه تارهای عضلانی کارگر قرار می گیرند. علاوه بر قطر کوچک این نوع تارها، از دیگر مشخصه های آنان می شود به فقدان لوله های T و شبکه سارکوپلاسمی در آنها اشاره نمود . پتانسیل آرامش غشایی در آنها پایین بوده ، سرعت اوج گیری از مرحله صفرآهسته است و سرعت هدایت الکتریکی آنها در مقایسه با تار های پورکنژ کندتر می باشد.
مسیر های آناتومیکی میانبر گره دهلیزی بطنی
مسیر های آناتومیکی متعددی وجود دارد که رسانش الکتریکی از دهلیز به بطن ها توسط آنها بصورت کنار گذر (میانبر)از گره دهلیزی بطنی محقق می گردد. تار های جیمزانواعی هستند که منشاء دهلیزی داشته ، از پیرامون گره دهلیزی بطنی عبور نموده و مجدداً از حاشیه بطنی آن وارد شده یا تا دیوراه بین بطنی رسوخ می نمایند. تار های ماهایم(نسبتاً اختصاص یافته ) کوتاه بوده ، ارتباطی مستقیم میان گره دهلیزی بطنی، تار های هیس یا شاخه های از آن نوار و دیواره بین بطنی را برقرار می کنند . نوار فرعی کنت که بافت های هدایتی اختصاص یافته یا میوکاردی سمت چپ و راست هستند ، پلی را میان دهلیز و بطن ایجاد می کنند. اگر چه نوار فوق غالباً در قلب جنین و نوزادان حضور دارد، ولی ممکن است در قلب برخی از بالغین نیز دیده شود. یاخته های پورگنژ در این نوار فرعی دیده نمی شوند.
تمامی مسیر های یاد شده بعنوان راههای هدایتی ممکنی در نظر گرفته می شوند که قادرند به هنگام فعال شدن، ناهنجاری هایی در رسانش گره دهلیزی بطنی را سبب گردند، بطوری که گره فوق را کنار گذاشته و به تکانه های قلبی که از دهلیز به بطن رسیده اند اجازه هدایت سریع را می دهند(بعنوان مثال پیش تحریکی بطنی) . مسیر های ناهنجاری که گره دهلیزی بطنی با رسانش آرام را بطور کامل دور می زنند(بعنوان مثال تارهای جیمز و نوار کنت) ممکن است موجب هدایت سریع تکانه های قلبی از دهلیز به بطن شده که آن هم سبب کوتاه شدن زمان تاخیر مابین تحریکات دهلیزی و بطنی از حد معمول خود می گردد(پیش تحریکی بطنی). الگوی الکتروکاردیوگرافی ناشی از این حالت (فاصلهPR کوتاه و QRS طویل) بخشی از نشانگان ولف- پارکینسون- سفید بوده که ممکن است با هم آوری بدآهنگی های سریع(تند بدآهنگی) بدلیل ورود مجدد جریانهای دخیل ، پیچیده تر گردد.
بطور خلاصه می شود گفت جریانات معمول فعالیت الکتریکی قلب به ترتیب ذیل می باش:
تکانه های قلبی از گره سینوسی منشاء می گیرند. قطبش وارون (فعالیت الکتریکی) از طریق یاخته های میوکارد دهلیز در جهت گره دهلیزی بطنی انتشار می یابند.آنگاه تکانه های فوق در حین گذر از بافت هدایتی اختصاص یافته در گره دهلیزی بطنی به ملایمت کند می شود.سرعت تکانه ها در طی عبور از نوار هیس، انشعابات نوار هیس و سامانه پورکنژ افزایش می یابد. تاخیر گره دهلیزی بطنی این اجازه را به دهلیز میدهد تا قبل از ورود خون به بطن ها انقباض خود را کامل نماید تا بدین واسطه با انقباض بطنی تداخل ننماید. تار های انتهایی پورکنژ با پایانه انشعابات نوار هیس اتصال یافته تا شبکه های درهم تافته بر روی سطح زیرین اندوکارد هر دو بطن شکل بگیرند که در نتیجه تکانه های قلبی تقریباً بصورت همزمان به عضلات قلبی بطن ها ی چپ و راست منتقل می شوند. تار های پورکنژ تمایل کمی برای انقباض در قاعده بطن و نوک عضلات پردی(پاپیلی) دارند. در عضله قلبی با فواصل متفاوتی نفوذ می کنند ، که بستگی به گونه حیوان دارد، در انسان و سگ تار های فوق تنها تا ثلث عضله قلبی وارد می شوند، در حالی که در سم داران تقریباً قادرند تا لایه اپی کارد برسند. آنگاه تکانه های عصبی از طریق یاخته های عضله قلبی بطن ها به طریق هدایت یاخته به یاخته انتشار می یابند.
قوانین پایه الکتروفیزیولوژی
هر یاخته قلبی دارای پتانسیل عمل غشایی مشخصی بوده (جدول 2-7 و تصویر 4-7)که مسئول انتقال تکانه های قلبی از یک یاخته به یاخته بعدی است. این پتانسیل های عمل غشایی متحرک ، در بارهای بیرونی سطح یاخته های عضلانی قلب تغییرات انتقالی بوجود می آورند. تفاوت های پتانسیلی متوالی که در عضله قلبی بوجود می آید یک میدان الکتریکی متفاوتی
جدول: 2-7: ویژگی های پتانسیل غشایی در قلب پستانداران.
را ایجاد می کند که به هنگام ثبت در سطح بدن از آن تحت عنوان الکتروکاردیوگرام (ECG) یاد می شود (تصویر 5-7).
تصویر 4-7:اشکال پتانسیل عمل غشایی بخش های مختلف قلب. دقت شود که تداوم پتانسیل عمل غشایی تا رسیدن به تار های پوکنژ طولانی و طولانی تر شده و در مورد عضلات بطنی طول این تداوم عمل کاهش می یابد . دلیل این پدیده سازو کاری است که به آن دروازه دار شدن(Gating) می گویند.
ویژگی های درون یاخته ای چون خودکاری، تحریک پذیری، قابلیت هدایتی و سرکشی مورد بحث قرار خواهند گرفت.این ویژگی ها به هنگام تجزیه و تحلیل بدآهنگی های قلبی وبا توجه به دارو های مخالف بدآهنگی ، از اهمیت برخوردار هستند .
تصویر 5-7: نمای بزرگ شده ای از الکتروکاردیوگرام اشتقاق دوم مجموعه P-QRS-T معمول در یک سگ به همراه فواصل زمانی و نشانه گذاری. اندازه گیری دامنه (برحسب میلی ولت) بواسطه حرکات مثبت و منفی نشان داده شده؛ فواصل زمانی (برحسب هزارم ثانیه) از سمت چپ به راست نشان داده شده. سرعت حرکت کاغذ نوار 50 میلیمتر در ثانیه؛1سانتی متر = یک میلی ولت).
پتانسیل عمل عرضی غشای یاخته های قلبی (تصویر 6-7 تا 8-7) مشتمل بر 5 مرحله به شرح ذیل می باشد:
- مرحله 1=باز قطبیدگی آغازین سریع
- مرحله 2= کفه
- مرحله3=بازقطبید گی سریع نهایی
- مرحله4=پتانسیل آرامش غشایی و قطبش وارون دیاستولی
تصویر 6-7: مراحل و شیب های پتانسیل عمل غشای یاخته های عضلانی قلب.
تصویر 8-7: پتاسیل های عمل. الف) یک تار عضلانی قلب . ب) یک تار پورکنژ. پیکان ها نقاطی هستند که الکترود های مویی به داخل یاخته وارد شده اند . توجه گردد که منحنی چنانکه پتانسیل غشایی ثبت می گردد بطور ناگهانی تا 90- میلی ولت تقعر می یابد. روند قطبش وارون دیاستولی (مرحله 4) در تار های پورکنژ بوجود می آید ولی در تارهای عضلانی قلب دیده نمی شود. تار های عضلانی در انتظار تحریک حاصل از پتاسیل عمل یاخته مجاور هستند.
تصویر 7-7:نموداری تصویری از انواع پتانسیل های عمل عرضی غشای. محور عرض ها بر مبنای میلی ولت . ERP= مرحله سرکشی موثر. دو پتانسیل عمل سمت راست به ترتیب مربوط به یاخته عضلانی قلب و یک یاخته پورکنژ هستند. این پتانسیل های عمل دارای دو سازه می باشند . یک پتانسیل کم پای تندو تیز(پاسخ سریع) و یک مرحله کفه ای آهسته تر(پاسخ آرام). یاخته های پاسخ سریع دارای یک قطبش وارون آغازین سریع هستند (مرحله 0)،متعاقب با یک باز قطبیدگی آغازین سریع (مرحله 1 ،عمدتاً در یاخته پورکنژ )، یک کفه (مرحله 2)، یک باز قطبیدگی نهایی (مرحله3) تا سطح پتانسیلی دیاستولی و یک پتانسیل دیاستولی (مرحله 4) هستند. مرحله 4 یک پتانسیل غشایی ثابت یا آرامش در یاخته های عضلانی قلب است ، اما در یاخته هایی از نوع پیش آهنگ (پورکنژ) تا رسیدن به پتانسیل آستانه ای تدریجاً قطبش وارون دیاستولی(مرحله4) محقق شده و پتانسیل عمل دیگری تولید می شود . در هر دو نوع از یاخته های پاسخ سریع پتانسیل سرکش موثر(ERP) کاهی در طی مرحله 3 به پایان می رسد . پتانسیل عمل در سمت راست پتانسیل عرضی غشای کند پاسخ گره سینوسی دهلیزی را نشان می دهد. مرحله 0 آهسته بوده، مرحله 1 حذف شده ، مرحله 3 تدریجی است، در طی مرحله 4 ممکن است یک نوسان پس پتانسیلی طی مرحله 4 پدیدار شده یا محقق نشود و مرحله 4 قطبش وارون ادامه یافته تا اینکه مرحله 0 دوباره آغاز گردد. پتانسیل سرکش موثر گره سینوسی دهلیزی ممکن است از طول مدت پتانسیل عمل فراتر رود.
مراحل فوق در نتیجه سیلان درونریز یونی که بصورت غیرفعال است محقق می شوند. یون ها در جهت نزولی شیب الکترو شیمیایی که با واسطه تلمبه سدیمی و سازوکارهای تبادلی تثبیت گردیده حرکت می نمایند. در آغاز هر یونی از طریق کانال یونی مختص خود می گذرد.تکانه ها بدون نیاز به ورودی عصبی (یکپارچگی عملیاتی ) توسط بخش هایی از یاخته با مقاومت الکتریکی کم از یاخته ای به یاخته بعدی اشاعه می یابند. فعالیت الکتریکی درون یاخته ای را می توان با واسطه دخول یک الکترود مویی با قطر راسی کمتر از 5/0 میکرومتر در یک یاخته منفرد اندازه گیری نمود. این الکترود مویی کمترین آسیب را به یاخته وارد می سازد،نقطه ورودی آن به سرعت فلسی می شود . 5 مرحله پتانسیل عمل یاخته قلبی مشتمل بر توالی سریع تغییرات در پتانسیل الکتریکی عرضی غشای یاخته ای است که در طی روند های قطبش وارون و باز قطبیدگی محقق می گردد . مرحله 4 دیاستول الکتریکی بوده و مرحله 0 تا 3 نشانگر سیستول الکتریکی است . در پایان مرحله 3 و آغاز مرحله 4 ، یاخته بطور کامل باز قطبیده شده و به بیشینه پتانسیل استراحتی خود می رسد. یاخته های غیر پیشآهنگ پتانسیل غشایی استراحت را تا رسیدن به پتانسیل آستانه ای با واسطه جریانی از قطبش وارون بافت مجاور، ثابت نگاه می دارند. در یاخته های پیشآهنگ مرحله 4 به خودی خود و به آرامی کاهش می یابد(به میزان کمی منفی می شود) . همچنین قطبش وارون مرحله 4 قطبش وارون دیاستولی آهسته یا خودکاری نامیده می شود . رخداد فوق روندی معمول در یاخته های پیشآهنگ می باشد ، اگر چه این رخداد ممکن است در دیگر یاخته های قلبی بطور مرضی محقق شود. در یاخته های پیشآهنگ پتانسیل غشای استراحت طی مرحله 4 کمی منفی شده تا اینکه پتانسیل آستانه فرارسیده یا اینکه با واسطه یک جریان خارجی (مانند دیگر پیشآهنگ قلبی ) یاخته به پتانسیل آستانه کشیده می شود. پتانسیل آستانه ای ولتاژی است که یاخته بایستی پیش از تحقق کامل قطبش وارون تا آن میزان کاهش یابد. بجز یاخته های پیشآهنگ که خود شان به آن آستانه می رسند ، دیگر یاخته های قلبی طی مرحله 4 در پتانسیل استراحتی خود مانده و با واسطه دریافت تکانه هدایتی از بافت قطبش وارون یافته مجاور به پتانسیل آستانه ای میرسند. مطالعات الکتروفیزیولوژی نشان می دهد که قطبش وارون دیاستولی به دلیل سیلان درون سوی(درون ریز) آهسته یون های سدیمی است (تصویر 9-7). مطالعات الکتروفیزیولوژیکی اخیر بر روی تار های پورکنژ نشان داده که جریان پیشآهنگ یک جریان درون ریز یون سدیمی (INa) است که در 50- میلی ولت غیر فعال بوده وبه آرامی با واسطه روند ازدیاد قطبیت فعال می شود . ضریب رسانایی پتاسیمی(gK+) متعاقب قطبش وارون افت نموده چرا که روند جبرانی درون ریزی افزایش می یابد. هنگامی که غشای یاخته قطبش وارون می یابد جریان درون ریزی آن ساده تر از جریان برون ریزی است . کاهش ضریب رسانش پتاسیمی به تولید قطبش وارون کمک می نماید . مرحله 0 فرازش مقدماتی و کلی پتانسیل عمل بوده و مرحله قطبش وارون سریع است که به هنگام نزدیک شدن یاخته قلبی به پتانسیل آستانه و در زمانی که کانال های تند سیلان سدیمی فعال شده اند، تحقق می یابد.
تصویر 9-7: جریان پیشآهنگ . طی مرحله 4 در یاخته های پیشآهنگ افزایش آهسته ای در میزان یونهای مثبت بوجود میآید(قطبش وارون آهسته دیاستولی) که ناشی از افزایش میزان ورود یون های سدیم به داخل یاخته است تااینکه یاخته به پتانسیل آستانه ای رسیده و به سرعت تغییر قطبیت می دهد. نشت برون یاخته ای پتاسیم (gK+)در طی این مرحله متفاوت است .
سرعت فرازش مرحله 0 (بعنوان مثال بیشینه سرعت (VMax) یا بیشینه تغییر ولتاژ در هر ثانیه) با تعداد یونهای سدیم قابل دسترسی و تعداد کانال های سدیمی باز تعیین می گردد، که متعاقباً بستگی به پتانسیل غشایی یاخته در وضعیت استراحت دارد ( تصاویر 10-7 و 11-7) .
تصویر 10-7: نمایش از کانال تند سیلان سدیمی(فعال شدن، غیر فعال شدن و بازگشت).
تصویر 11-7:نمایی از کانال سدیمی کلسیمی کند سیلان(فعال شدن ، غیرفعال شدن و بازگشت).
در طی مرحله 0 پتانسیل عمل تند پاسخ ، جریان درون ریز ثانویه ای در پایین ترین حد پتانسیل غشایی آغاز می گردد. این جریان ثانویه توسط یونهای کلسیم و سدیم منتقل شده ، سرعت فرازش آن (Vmax) و سرعتی که با آن به دیگر یاخته های انتشار می یابد خیلی آهسته تر از آنی است که توسط جریان تند سدیمی تولید می شود . این جریان آهسته مسئول قطبش وارون مرحله 0 در یاخته هایی است که فاقد کانال های سدیمی تند سیلان بوده (مانند؛ یاخته های گره سینوسی و یاخته های گره دهلیزی بطنی ) و آنهایی که تنها کانال های کلسیمی کند سیلان دارند.
مرحله 1 شروع سریع و کوتاهی است از روند باز قطبیدگی به فوریت پس از مرحله 0 . این مرحله به سبب بسته شدن ناگهانی کانال های سدیمی تند سیلان و جریان پتاسیمی برون ریز گذرا پدیدار می گرد. مرحله 1 مختوم به کفه پتانسیل عمل است .
مرحله 2 (کفه) با واسطه روند درون ریزی یون های مثبت(کلسیم و سدیم) از طریق کانال های کند سیلان کلسیمی بداخل یاخته تثبیت می گردد(تصاویر 6-7 و 11-7).این مرحله مرحله کفه پتانسیل عمل بوده که در حکم مرحله نسبتاً طولانی سرکشی در یاخته های عضلانی قلب در مقایسه با یاخته های عضلات اسکلتی است (تصویر 12-7).
تصویر 12-7: الف) دامنه و سرعت بیشینه (Vmax) در ارتباط با پتانسیل غشایی هستند که از آن برخاسته اند. پاسخ های اولیه (a,b) پتانسیل عمل کند پاسخ هستند که از پتانسیل غشایی نسبتاً مثبت منشاء می گیرند، این پاسخ ها گسترش نمی یابند. پاسخ نوع C زود هنگام ترین انتشار پتانسیل عمل است (پاسخ سریع فروکش می کند)، اما بدلیل پایین بودن سرعت بیشینه و دامنه آن به آهستگی گسترش می یابند. پاسخ نوع d قبل از کاهش روند باز قطبیدگی بر انگیخته شده و دارای بیشینه سرعت و دامنه بزرگتری در مقایسه با نوع C می باشد، زیرا که از پتانسیل منفی تری نشات می گیرد. این نوع پاسخ نیز کند تر از حالت معمول گسترش می باشد . پاسخ نوع e پس از کامل شدن روند باز قطبیدگی که غالباً دارای پتانسیل آرامش منفی است، دارای نسبتdv/dt (شتاب) و دامنه و سرعت انتشار معمول می باشد . TP=پتانسیل آستانه ای؛ RP= پتانسیل آرامش.
ب) ARP=طول تخمینی دوره سرکشی مطلق؛ TRP=کل دوره سرکشی؛ ERP=دوره سرکشی موثر، FRT = کل زمان باز یابی؛ SNP=دوره پیش از وضعیت عادی؛ RRP دوره سرکشی نسبی. پاسخ هایی که بصورت خطوط نقطه چین نشانداده شده اند، a وb در شکل الف در شمار پاسخهایی طبقه بندی می شوند که انتشار ندارند. پاسخ نوع C زود هنگام ترین پاسخ قابل انتشار بوده و اختتام مرحله سرکشی موثر را تعیین می نماید. پاسخ فوق پس از پایان روند باز قطبیدگی بر انگیخته شده و ازدیاد سرعت و دامنه آن در حد معمول بوده ، پایان مدت باز گشت کامل را تعیین می نماید. تغییرات در میزان آستانه ای که در شکل ب نشان داده شده در ارتنباط با قدر مطلق شدت جریان می باشد. منحنی آغاز همزمانی عدم تحریک پذیری با مرحله 0 پتانسیل عمل غشایی ، کاهش تدریجی آستانه در طی مرحله 3 روند بازقطبیدگی و قابلیت مجدد تحریک ذیری کامل پس از اختتام مرحله 3 پتانسیل فعالیت غشایی را نشان می دهد( مرحل در تصویر 10-7 معین شده است) .
این جریان کلسیمی درون ریز کند یاخته را در وضعیت قطبش وارون طولانی نگاه داشته ، تا پیش از آن که تکانه ای دیگر آغاز گردد زمان کافی برای تحقق انقباض عضلانی مهیا باشد . مرحله 2 برای انقباض عضله قلبی حائز اهمیت است چرا که دخول یون های کلسیم به افزایش غلظت کلسیم سیتوپلاسمی یاخته عضلانی که لازمه همبست تحریک- انقباض است کمک می نماید. همچنین این افزایش کلسیم درون یاخته ای سبب رهایش بیشتر کلسیم های اضافی از شبکه سارکوپلاسمیک می شود . درونریزی آهسته سدیم و جریانات کلسیمی ، همچنین برون ریزی (خروج) جریان یون های کلر از یاخته ، مرحله کفه ای مثبت پتانسیل غشایی را تثبیت می نماید.
مرحله 3 بازقطبیدگی سریع پایانی پتانسیل عمل است. این مرحله ناشی از غیرفعال شدن کانال های درون ریز کند سیلان کلسیمی ( در ادامه بحث می آید)، و فعال شدن کانال های برونریز پتاسیمی است . فعالیت تلمبه سدیمی- پتاسیمی با واسطه غلظت بالای سدیم درون یاخته ای افزایش یافته و وظیفه آن ذخیره غلظت سدیم و پتاسیم تا سطح استراحت آنها است . تلمبه فوق برق زا(مولد الکتریکی) بوده و سهم کوچکی (10 میلی ولت) در پیدایش پتانسیل عرضی غشای دارد.
حداقل دو کانال پتاسیمی وابسته به ولتاژ در این خصوص دخالت دارند. یکی از این کانال ها به پتاسیم اجازه نشت درون ریزی در مقابل شیب الکتروشیمیای پتاسیم می دهد(چنانکه به آن درونرریزی یکسویه یا یکسو سازی ناهنجار می گویند) . این کانال مسئول کاهش سریع رسانش پتاسیمی در قطبش وارون (طی مرحله 2) و افزایش رسانش با باز قطبیدگی است. دومین کانال پتاسیمی اجازه عبور برون ریز به یون ها را می دهد(در جهت نزولی شیب الکتروشیمیایی پتاسیم). فعال شدن آن موجب افزایش رسانش کلی پتاسیمی شده که کفه مرحله 2 را مختوم می نماید.
Transmembrane action potential

